

European Journal of Neurodegenerative Diseases 2019; 8(2) July-December 22-30
EXPERIMENTAL STUDY ON VASOCONSTRICTION AND INFLAMMATION: ROLE OF LIPOXYGENASE PRODUCTS
G.Neri1* and R.G. Bellomo2
1Department of Neurosciences, Imaging and Clinical Sciences, University G. d’Annunzio Chieti-Pescara, Chieti, Italy
2Department of Biomolecular Sciences, University of Study of Urbino ‘Carlo Bo’, Urbino, Italy
*Correspondence to:
Giampiero Neri, MD
Department of Neurosciences, Imaging and Clinical Sciences,
University G. d’Annunzio Chieti-Pescara,
Chieti, Italy
e-mail: giampiero.neri@unich.it
| Received: 22 September, 2019 Accepted: 27 October, 2019  |
2279-5855 (2019) Copyright © by BIOLIFE This publication and/or article is for individual use only and may not be further reproduced without written permission from the copyright holder. Unauthorized reproduction may result in financial and other penalties Disclosure: all authors report no conflicts of interest relevant to this article. |
ABSTRACT
A sufficient blood supply is vital for the physiological functioning of the brain, and a lack of cerebral blood flow leads to neurodegenerative diseases. Immune cells defend the brain system, but they can be protagonists of inflammatory processes in pathological cases. Vasoconstriction and inflammation are pathological elements of the brain. Here, we study the pathogenetic mechanisms of vasoconstriction and inflammation in a rodent model in relation to arachidonic acid compounds. Experimentally induced inflammation was treated with various lipoxygenase and cyclooxygenase inhibitors. Lipoxygenase was inhibited by specific compounds but not by cyclooxygenase inhibitors, although both were anti-inflammatory.
KEYWORDS: lipoxygenase, arachidonic acid, leukotriene, hydroxyeicosatetraenoic acids, leukocytes, SRS-A
INTRODUCTION
The brain requires a sufficient blood supply for physiological functioning. A deficiency of cerebral blood flow leads to neurodegenerative diseases (1). In recent years, the importance of lipoxygenase as a key enzyme in inflammatory reactions and allergic processes has been increasingly recognised. The metabolites of arachidonic acid generated after stimulation of the lipoxygenase pathway (2), e.g., the hydroxy fatty acids and the leukotrienes, can regulate both cellular and humoral components of inflammatory and allergic reactions (6). In the first figure, some activities of lipoxygenase products are shown (Fig. 1).
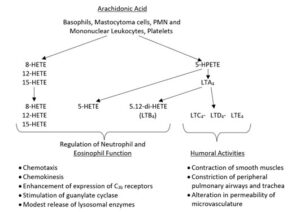
Fig. 1. Generation and biological properties of products of arachidonic acid. HPETE = hydroperoxyeicosatetraenoic acid, LT = leukotriene.
The hydroxyeicosatetraenoic acids (HETEs) possess leukotactic and leukinetic activities in which the potencies differ from the kind of metabolite (3). Leukotriene B4 is the most potent chemotactic factor for polymorphonuclear leukocytes. Vascular effects of this metabolite have also been observed due to increased permeability (4).
The stimulation of the leukocyte migration and the vascular effects are reactions which are important for the responses during inflammatory processes (5). Remarkably, some lipoxygenase products have modulating influences on the immune system (6). Thus, the expression of C3b receptors is enhanced, and various lipoxygenase products induce T-suppressor cells. Another important action for inflammatory and allergic reactions is the release of lysosomal enzymes (7).
The mechanism of action of the lipoxygenase products is possibly mediated via stimulating the guanylate cyclase and increasing the cGMP level (8). The leukotrienes C4, D4, and E4, components of SRS-A, are mediators released mainly during allergic reactions while also having been reported to play a role in inflammation (9). They have humoral activities and contract specific smooth muscles, e.g., ileum, peripheral pulmonary airways, and trachea. In addition, these compounds increase vascular permeability, leading to plasma exudation (10, 11).
Due to the multiple actions of the lipoxygenase products, potent lipoxygenase inhibitors with favourable pharmacokinetics should, therefore, influence inflammatory and allergic reactions. Combining such agents with antagonists of other mediators of inflammation and allergic reactions could lead to more successful treatment (12).
The study of the pharmacological properties of drugs influencing inflammation and allergy is complicated by several different mediators that can be released during these reactions (13). It is further complicated by the fact that some of these mediators can have opposite effects and by the variety of animal models employed, some of which may not simulate the pathological situation in man (14).
An optimal test hierarchy should include investigating drugs’ influence on the isolated enzyme, and such models in which mediators synthesised by the lipoxygenase are important for the pathological process (15).
We tested some compounds (Table I) to inhibit the reticulocyte lipoxygenase from rabbit and soybean lipoxygenase 1.
Table I: Investigated compounds.
| Investigated compounds |
| BW 755 C |
| Nordihydroguaiaretic acid (NDGA) |
| Salicylhydroxamic acid (SHAM) |
| FPL 55712 |
| Propyl gallate |
| Benoxaprofen |
| Diclofenac -Na |
| Indomethacin |
| Acetylsalicylic acid (ASA) |
| Phenylbutazone |
| Noradrenaline |
| Adrenaline |
| Isoprenaline |
| H 252 |
| M 516 |
The reticulocyte lipoxygenase converts arachidonic acid into the major product 15-HETE, and 12-HETE as the minor component (16), and the soybean lipoxygenase-1 forms 15-HPETE from arachidonic acid (17).
The anti-inflammatory and antiallergic activities of the compounds have been investigated by using the carrageenin oedema of rat paw as a model of acute inflammation, the adjuvant arthritis of the rat as a model of chronic inflammation, and the active anaphylactic oedema of rat paw as well as the active cutaneous anaphylaxis of the rat, as allergic models (18).
MATERIALS AND METHODS
Reticulocyte lipoxygenase was isolated from the defibrinated blood of rabbits and made anaemic by repeated bleeding or treatment with phenylhydrazine. The purification procedure involved ammonium sulfate precipitation, anion exchange chromatography on DEAE-Sephadex A-50, and isoelectric focusing in ampholine, pH 5 to 7.
Soybean lipoxygenase was obtained from Boehringer Mannheim GmbH (FRG). The activity of the lipoxygenase was measured polarographically through an oxygen electrode. The reaction mixture contained 0.53 mM linoleic acid, 0.2% sodium cholate, and 5% ethanol in 0.1 M potassium phosphate buffer, pH 7.4 and 9.0, respectively, and the final volume was 2.0 ml. Inhibitors were preincubated with the enzyme for 10 min. The assays were performed at 20° C.
Carrageenin oedema of the rat paw was induced in female Wistar rats by subplantar injection of 0.1 ml of a 1% carrageenin solution. The drugs were given orally and simultaneously with the carrageenin injection. Adjuvant arthritis was induced in female Wistar rats by subplantar injection of 0.1 ml Freund’s complete adjuvant (0.5% suspension of heat-killed mycobacterium. in paraffinic perliquidum). The drugs were given orally once a day.
For producing the anaphylactic reactions, female Wistar rats were sensitised by i.m. injecting 0.2 ml of a 0.5% solution of bovine serum albumin in Bordetella pertussis vaccine containing 2×1010 bacteria. Two weeks later, the animals were challenged by subplantar injection of 500 mg bovine serum albumin in 0.1 ml 0.9% NaCl solution for producing the active anaphylactic oedema and by intracutaneous injection of 0.5 mg bovine serum albumin in 0.05 ml 0.9% NaCl solution for performing active cutaneous anaphylaxis, respectively. The drugs were administered orally 1 hour before the provocation of the edema and intraperitoneally 20 minutes before the cutaneous anaphylaxis, respectively.
RESULTS AND DISCUSSION
Table II. Inhibition of the activity of the reticulocyte lipoxygenase and soybean lipoxygenase-1.
| Compound | Inhibitory potency
Reticulocyte lipoxygenase
|
Soybean
lipoxygenase-1 |
|
| BW 755 C | IC50= 3×10-6 M/l | IC50= 1.7×10-4 M/l | |
| NDGA | IC50= 4×10-6 M/l | IC50= 2×10-5 M/l | |
| SHAM | IC50= 5.6×10-5 M/l | IC50= 4.5×10-4 M/l | |
| FPL 55712 | IC50= 4×10-4 M/l | 10-3M/l 39% inhib. | |
| Propyl gallate | IC50= 4×10-5 M/l | IC50= 9.4×10-5 M/l | |
| Benoxaprofen | 10-3M/l 50% inhib. | 10-3M/l no inhib. | |
| Diclofenac | 10-3M/l 53% inhib. | 10-3M/l no inhib. | |
| Indomethacin | 10-3M/l 51% inhib. | 10-3M/l no inhib. | |
| ASA | 10-3M/l no inhib. | 10-3M/l no inhib. | |
| Phenylbutazone | 10-3M/l no inhib. | 10-3M/l no inhib. | |
| Adrenaline | 10-3M/l no inhib. | 10-3M/l no inhib. | |
| Noradrenaline | 10-3M/l 27% inhib. | 10-3M/l no inhib. | |
| Isoprenaline | 10-3M/l no inhib. | 10-3M/l no inhib. | |
| H 252 | 10-3M/l no inhib. | 10-3M/l no inhib. | |
| M 516 | IC50= 6×10-5 M/l | IC50= 5.4×10-4 M/l |
Generally, soybean lipoxygenase seems more insensitive to the inhibitors than reticulocyte lipoxygenase (19). Therefore, the most potent inhibitors are BW 755 C, inhibiting cyclooxygenase (15), NDGA, SHAM, and propyl gallate.
Remarkably, the SRS-A antagonist FPL 55712 inhibits the lipoxygenase activity, too and does not only display its antiallergic activity by antagonising the actions of the leukotrienes at the receptor site. The antiallergic and anti-inflammatory agent benoxaprofen blocks the lipoxygenase activity very weakly in our system (20). This drug selectively inhibits the 5-lipoxygenase, which is responsible for the biosynthesis of the precursors of LTC4, D4, and E4 (21).
Of the anti-inflammatory drugs, only diclofenac and indomethacin exert weak inhibitory activities against the reticulocyte lipoxygenase. Catecholamines inhibit inflammatory responses (22) but do not affect lipoxygenase activity. The compound M 516 is shown to be a strong, selective inhibitor of lipoxygenase because it does not inhibit cyclooxygenase (23). In investigating the compounds in the in vivo models, BW 755 C inhibits the carrageenin oedema after systemic and local administration (Tables III, IV).
Table III. Inhibition of the carrageenin edema of rat paw ( ++);( +) significantly different from control group (p<0.01; p<0.05) according to Student’s t-test.
| Compound
Dose/Administr. |
% Inhibition | |||||
| 0.5 h | 1 h | 2 h | 3 h | 4 h | 5 h | |
|
BW 755 C 50 mg/kg p.o. |
35++ |
37++ |
62++ |
62++ |
60++ |
|
| BW 755 C
1 mg local. |
29+ | 33+ | 21+ | -20 | -10 | |
| NDGA
200 mg/kg p.o. |
0 | 0 | 0 | 0 | 0 | |
| SHAM
200 mg/kg p.o. |
9 | 0 | 0 | 0 | 7 | |
| FPL 55712
0.2 mg local. |
45++ | 25 | -37 | -4 | -7 | 5 |
| Propyl gallate
250 mg/kg p.o. |
39++ | 19 | ||||
| Propyl gallate
1 mg local. |
0 | 20 | 9 | |||
| Benoxaprofen
20 mg/kg p.o. |
41+ | 35+ | 23 | 16 | 19 | |
| Diclofenac
2.5 mg/kg p.o. |
49++ | 22 | ||||
| Indomethacin
0.1 mg/kg p.o. |
35++ | 41++ | 34++ | |||
| Indomethacin
0.1 mg local. |
38++ | 49++ | 45++ | |||
| ASA
250 mg/kg p.o. |
53++ | 57++ | 53++ | |||
| Phenylbutazone
100 mg/kg p.o. |
57++ | 62++ | 54++ | |||
| Phenylbutazone
0.5 mg local. |
38+ | 57+ | 45+ | 38+ | 22+ | |
| Adrenaline
0.01 ug local. |
49++ | 54++ | 7 | 9 | 17 | |
| Noradrenaline
0.01 ug local. |
27+ | 18 | 13 | 4 | 6 | |
| Isoprenaline
0.01 ug local. |
29+ | 36+ | 10 | 7 | 7 | |
| H 252
65 mg/kg p.o. |
58++ | 50++ | 26 | 7 | ||
| M 516
68 mg/kg p.o. |
41+ | 45+ | 45+ | 34 | ||
| M 516
1 mg local.
|
37+ | 49+ | 0 | -18 | 2 | |
Table IV. Inhibition of the adjuvant arthritis of rat ( ++);( +) significantly different from the control group (p<0.01; p<0.05) according to Student’s t-test.
| Compound
Dose/Administr. |
% Inhibition | ||
| 2 d | 4 d | 16 d | |
|
BW 755 C 50 mg/kg p.o. |
23+ |
20+ |
|
| Propyl gallate
250 mg/kg p.o. |
2 | 6 | 0 |
| Benoxaprofen
20 mg/kg p.o. |
33+ | 35+ | |
| Diclofenac
2.5 mg/kg p.o. |
41++ | 51++ | 52++ |
| Indomethacin
1 mg/kg p.o. |
25+ | 42++ | 14+ |
| ASA
250 mg/kg p.o. |
44++ | 49++ | |
| Phenylbutazone
100 mg/kg p.o. |
52++ | 59++ | |
| Adrenaline
0.25 mg/kg s.c. |
9 | 0 | |
| M 516
68 mg/kg p.o.
|
0 | 0 | |
Strong inhibitory properties on the anaphylactic models are also evident, as shown in Tables V and VI. These results conclude that there is the best correlation between in vitro and in vivo activities. However, the in vivo effects can also be caused by inhibiting cyclooxygenase.
Table V. Inhibition of the active anaphylactic oedema of rat paw (+); significantly different from the control group (p<0.05) according to Student’s t-test.
| Compound
Dose/Administr. |
% Inhibition | |||||
| 0.5 h | 1 h | 2 h | 3 h | 4 h | 5 h | |
|
BW 755 C 50 mg/kg p.o. |
27 |
29 |
40+ |
38+ |
34+ |
41+ |
| Benoxaprofen
50 mg/kg p.o. |
42+ | 47+ | 51+ | 38+ | 54+ | 52+ |
| Diclofenac
3 mg/kg p.o. |
-7 | 0 | 15 | 20 | 14 | 23 |
| Indomethacin
5 mg/kg p.o. |
14 | 16 | 25 | 20 | 25 | 28 |
| ASA
200 mg/kg p.o. |
12 | 32 | 35+ | 27 | 52+ | 31 |
| Phenylbutazone
30 mg/kg p.o. |
18 | 7 | 2 | 18 | 5 | 27 |
| H 252
100 mg/kg p.o. |
14 | 27 | 13 | 4 | 6 | 8 |
| M 516
100 mg/kg p.o.
|
6 | 3 | 24 | 2 | -3 | 1 |
Table VI. Inhibition of the active cutaneous anaphylaxis of the rat (+); significantly different from the control group (p<0.05) according to Student’s t-test.
| Compound
Dose/Administr. |
% Inhibition |
|
BW 755 C 50 mg/kg i.p. |
43+ |
| Benoxaprofen
50 mg/kg i.p. |
48+ |
| Diclofenac
5 mg/kg i.p. |
6 |
| Indomethacin
5 mg/kg i.p. |
22 |
| Phenylbutazone
30 mg/kg i.p. |
12 |
| ASA
200 mg/kg i.p. |
27 |
| H 252
100 mg/kg i.p. |
22 |
| M 516
100 mg/kg i.p.
|
18 |
The antioxidant propyl gallate inhibits carrageenin oedema only after oral administration at relatively high doses. Therefore, it does not affect adjuvant arthritis (24).
SHAM and NDGA show no influence on carrageenin oedema. Despite the suitable inhibitory activities on the lipoxygenase, there is no correlation with the in vivo properties of the propyl gallate, SHAM, and NDGA compounds. The cause of this infectivity could be insufficient absorption with ineffective serum and tissue concentration. The SRS-A antagonist FPL 55712 decreases the swelling of the carrageenin oedema in the first hour, suggesting that the C4, D4, and E4 leukotrienes or other lipoxygenase products are released (25).
As an inhibitor of the 5-lipoxygenase, benoxaprofen acts dose-dependently on carrageenin oedema and adjuvant arthritis; these results are also obtained in the anaphylactic models. Acid nonsteroidal anti-inflammatory agents have the strongest inhibitory activities on carrageenin oedema and adjuvant arthritis and are cyclooxygenase inhibitors (26), while acetylsalicylic acid also significantly inhibits the anaphylactic paw oedema but not cutaneous anaphylaxis. Carrageenin oedema is inhibited by the catecholamines after local administration; however, inhibition of adjuvant arthritis could not be found (27).
The compounds H 252, which does not inhibit lipoxygenase activity, and M516, which inhibits lipoxygenase, show moderate dose-dependent inhibitory activities in carrageenin oedema. However, both drugs have no statistically significant influence on the anaphylactic models. The compound M 516 is also ineffective in the primary phase of adjuvant arthritis.
It can be stated that there are no satisfactory correlations between the inhibition of the 15- lipoxygenase and in vivo activities. The cause could be the subordinate importance of the products of these lipoxygenases in our in vivo models and/or insufficient substance concentrations in vivo. The best correlation is obtained with BW 755 C, which influences all in vivo models but is also a potent cyclooxygenase inhibitor.
CONCLUSION
The importance of lipoxygenase in inflammatory and allergic reactions is established. It oxidises multiple unsaturated fatty acids into many biologically active compounds, such as hydroxyeicosatetraenoic acids and leukotrienes. Therefore, the inhibitors of the lipoxygenase may be possible anti-inflammatory and antiallergic drugs (28).
Here, some compounds were tested for their activity against the isolated lipoxygenase from rabbit reticulocytes and soybean and in four in vivo models. Again, conventional anti-inflammatory drugs with high potency are tested as a standard.
The most potent lipoxygenase inhibitors were BW 755 C, NDGA, SHAM, and the compound M 516, which showed anti-inflammatory and antiallergic activities in some in vivo models (29). On the other hand, the potent cyclooxygenase inhibitors’ anti-inflammatory agents had no inhibitory action on the lipoxygenase, but they strongly blocked the inflammatory reactions in vivo (30).
Conflict of interest
The author declares that they have no conflict of interest.
REFERENCES
- Duvall MG, Levy BD. DHA- and EPA-derived resolvins, protectins, and maresins in airway inflammation. European Journal of Pharmacology. 2016;785:144-155. doi:https://doi.org/10.1016/j.ejphar.2015.11.001
- Powell WS, Rokach J. Biosynthesis, biological effects, and receptors of hydroxyeicosatetraenoic acids (HETEs) and oxoeicosatetraenoic acids (oxo-ETEs) derived from arachidonic acid. Biochimica et Biophysica Acta (BBA) – Molecular and Cell Biology of Lipids. 2015;1851(4):340-355. doi:https://doi.org/10.1016/j.bbalip.2014.10.008
- Bitencourt CS, Bessi VL, Huynh DN, et al. Cooperative role of endogenous leucotrienes and platelet‐activating factor in ischaemia–reperfusion‐mediated tissue injury. Journal of Cellular and Molecular Medicine. 2013;17(12):1554-1565. doi:https://doi.org/10.1111/jcmm.12118
- Byrum RS, Goulet JL, Snouwaert JN, Griffiths RJ, Koller BH. Determination of the Contribution of Cysteinyl Leukotrienes and Leukotriene B4 in Acute Inflammatory Responses Using 5-Lipoxygenase- and Leukotriene A4 Hydrolase-Deficient Mice. The Journal of Immunology. 1999;163(12):6810-6819. doi:https://doi.org/10.4049/jimmunol.163.12.6810
- Muller WA. Getting Leukocytes to the Site of Inflammation. Veterinary Pathology. 2013;50(1):7-22. doi:https://doi.org/10.1177/0300985812469883
- Catanzaro M, Corsini E, Rosini M, Racchi M, Lanni C. Immunomodulators Inspired by Nature: A Review on Curcumin and Echinacea. Molecules. 2018;23(11):2778. doi:https://doi.org/10.3390/molecules23112778
- Ribeiro D, Freitas M, Lima JLFC, Fernandes E. Proinflammatory Pathways: The Modulation by Flavonoids. Medicinal Research Reviews. 2015;35(5):877-936. doi:https://doi.org/10.1002/med.21347
- Dobrian AD, Lieb DC, Cole BK, Taylor-Fishwick DA, Chakrabarti SK, Nadler JL. Functional and pathological roles of the 12- and 15-lipoxygenases. Progress in Lipid Research. 2011;50(1):115-131. doi:https://doi.org/10.1016/j.plipres.2010.10.005
- Bäck M. Leukotriene Signaling in Atherosclerosis and Ischemia. Cardiovascular Drugs and Therapy. 2008;23(1):41-48. doi:https://doi.org/10.1007/s10557-008-6140-9
- Claesson-Welsh L. Vascular permeability—the essentials. Upsala Journal of Medical Sciences. 2015;120(3):135-143. doi:https://doi.org/10.3109/03009734.2015.1064501
- Nagy JA, Benjamin L, Zeng H, Dvorak AM, Dvorak HF. Vascular permeability, vascular hyperpermeability and angiogenesis. Angiogenesis. 2008;11(2):109-119. doi:https://doi.org/10.1007/s10456-008-9099-z
- Sala A, Proschak E, Steinhilber D, Rovati GE. Two-pronged approach to anti-inflammatory therapy through the modulation of the arachidonic acid cascade. Biochemical Pharmacology. 2018;158:161-173. doi:https://doi.org/10.1016/j.bcp.2018.10.007
- Gómez E, Torres MJ, Mayorga C, Blanca M. Immunologic Evaluation of Drug Allergy. Allergy, Asthma & Immunology Research. 2012;4(5):251. doi:https://doi.org/10.4168/aair.2012.4.5.251
- Gregory NS, Harris AL, Robinson CR, Dougherty PM, Fuchs PN, Sluka KA. An Overview of Animal Models of Pain: Disease Models and Outcome Measures. The Journal of Pain. 2013;14(11):1255-1269. doi:https://doi.org/10.1016/j.jpain.2013.06.008
- Meirer K, Steinhilber D, Proschak E. Inhibitors of the Arachidonic Acid Cascade: Interfering with Multiple Pathways. Basic & Clinical Pharmacology & Toxicology. 2013;114(1):83-91. doi:https://doi.org/10.1111/bcpt.12134
- Kim BJ, Shin KC, Oh DK. Enzymatic Production of 15-Hydroxyeicosatetraenoic Acid from Arachidonic Acid by Using Soybean Lipoxygenase. Journal of Microbiology and Biotechnology. 2014;24(3):359-362. doi:https://doi.org/10.4014/jmb.1310.10054
- Chawengsub Y, Gauthier KM, Campbell WB. Role of arachidonic acid lipoxygenase metabolites in the regulation of vascular tone. American Journal of Physiology-Heart and Circulatory Physiology. 2009;297(2):H495-H507. doi:https://doi.org/10.1152/ajpheart.00349.2009
- Fehrenbacher JC, Vasko MR, Duarte DB. Models of Inflammation: Carrageenan- or Complete Freund’s Adjuvant-Induced Edema and Hypersensitivity in the Rat. Current protocols in pharmacology / editorial board, SJ Enna (editor-in-chief) [et al]. 2012;0 5:Unit5.4. doi:https://doi.org/10.1002/0471141755.ph0504s56
- Youn B, Sellhorn GE, Mirchel RJ, Gaffney BJ, Grimes HD, Kang C. Crystal structures of vegetative soybean lipoxygenase VLX-B and VLX-D, and comparisons with seed isoforms LOX-1 and LOX-3. Proteins: Structure, Function, and Bioinformatics. 2006;65(4):1008-1020. doi:https://doi.org/10.1002/prot.21182
- Dawson W, Boot JR, Harvey J, Walker JR. The pharmacology of benoxaprofen with particular to effects on lipoxygenase product formation. European Journal of Rheumatology and Inflammation. 1982;5(2):61-68.
- Kobayashi I, Hamasaki Y, Sato R, et al. Saiboku-To, a herbal extract mixture, selectively inhibits 5-lipoxygenase activity in leukotriene synthesis in rat basophilic leukemia-1 cells. Journal of Ethnopharmacology. 1995;48(1):33-41. doi:https://doi.org/10.1016/0378-8741(95)01280-q
- Gan TJ. Diclofenac: an update on its mechanism of action and safety profile. Current Medical Research and Opinion. 2010;26(7):1715-1731. doi:https://doi.org/10.1185/03007995.2010.486301
- Rao CV, Janakiram NB, Mohammed A. Lipoxygenase and Cyclooxygenase Pathways and Colorectal Cancer Prevention. Current Colorectal Cancer Reports. 2012;8(4):316-324. doi:https://doi.org/10.1007/s11888-012-0146-1
- Woode E, Ainooson G, Amo-Barimah A, Boakye-Gyasi E, Abotsi WonderMK. Antiarthritic and antioxidant effects of the leaf extract ofFicus exasperataP. Beauv. (Moraceae). Pharmacognosy Research. 2010;2(2):89. doi:https://doi.org/10.4103/0974-8490.62958
- HSU HC, LIN WC, CHANG PJ, HONG CZ, CHEN CH. Propyl gallate inhibits TPA-induced inflammation via the nuclear factor-κB pathway in human THP-1 monocytes. Experimental and Therapeutic Medicine. 2013;5(3):964-968. doi:https://doi.org/10.3892/etm.2013.896
- Hiermann A, Schramm HW, Laufer S. Anti-inflammatory activity of myricetin-3-O-β-D-glucuronide and related compounds. Inflammation Research. 1998;47(11):421-427. doi:https://doi.org/10.1007/s000110050355
- Poh J, Knowles S. Safety of 5-Aminosalicylic Acid Derivatives in Patients with Sensitivity to Acetylsalicylic Acid and Nonsteroidal Anti-inflammatory Drugs. The Canadian Journal of Hospital Pharmacy. 2014;67(1). doi:https://doi.org/10.4212/cjhp.v67i1.1318
- Wisastra R, Dekker F. Inflammation, Cancer and Oxidative Lipoxygenase Activity are Intimately Linked. Cancers. 2014;6(3):1500-1521. doi:https://doi.org/10.3390/cancers6031500
- Batt DG. 1 5-Lipoxygenase Inhibitors and their Anti-inflammatory Activities. Progress in Medicinal Chemistry. 1992;29:1-63. doi:https://doi.org/10.1016/s0079-6468(08)70004-3
- Zarghi A, Arfaei S. Selective COX-2 Inhibitors: A Review of Their Structure-Activity Relationships. Iranian journal of pharmaceutical research : IJPR. 2011;10(4):655-683.