

European Journal of Neurodegenerative Diseases 2019; 8(2) July-December 18-21
HAEMORRHAGE AND STROKE: IMMUNITY AND INFLAMMATION
F. Carinci
Department of Translational Medicine, University of Ferrara, Ferrara, Italy
Correspondence to:
Prof. Francesco Carinci
Department of Translational Medicine,
University of Ferrara,
Ferrara, Italy
e-mail: crc@unife.it
| Received: 23 May 2019 Accepted: 07July 2019  |
2279-5855 (2019) Copyright © by BIOLIFE This publication and/or article is for individual use only and may not be further reproduced without written permission from the copyright holder. Unauthorized reproduction may result in financial and other penalties Disclosure: all authors report no conflicts of interest relevant to this article. |
ABSTRACT
In ischemic stroke, microglia and the blood-brain barrier (BBB) are activated. This phenomenon leads to the activation of immune cells such as monocytes/macrophages, lymphocytes, mast cells (MCs), and neutrophils. The recruitment of these cells to the site of tissue damage leads to the release of pro-inflammatory cytokines, while in normal tissue, these cells are beneficial for the brain’s physiological activity. The BBB protects brain tissue and can be disrupted after stroke, an effect mediated by pro-inflammatory cytokines. However, the relationship between the BBB, microglia and pro-inflammatory compounds in hemorrhagic stroke still needs to be determined.
KEYWORDS: haemorrhage, stroke, immunity, inflammation, cytokines, microglia, BBB
INTRODUCTION
Ischemic stroke is characterized by decreased blood flow in the brain tissue and the activation of microglia, which are macrophage-like immune cells resident in the central nervous system (CNS) (1). Microglia are brain cells adjacent to the microvasculature in the basal ganglia, substantia nigra, and hippocampus. These cells, like activated peripheral blood monocytes, produce inflammatory cytokines and chemokines but can also release activated mast cells (MCs), producing proteases and reactive oxygen species (ROS) (2). The release of these pro-inflammatory compounds in stroke recruits immune cells to the site and exacerbates brain damage, resulting in neuronal destruction (3). In addition, the recruitment of immune cells from the bloodstream into the damaged CNS leads to the upregulation of adhesion molecules and cytokine/chemokine receptors with traumatic brain injury (4).
Conversely, the arrival of immune cells occludes vessels after ischemia, causing neuroprotection (5). These cellular dynamics are still unclear and can be the subject of future studies. Ischemic brain tissue produces damage-associated molecular patterns (DAMPs) with immune and inflammatory responses contributing to stroke (6).
DISCUSSION
Immune cells, including T cells, macrophages, microglia, MCs, and neutrophils, are present after stroke and cause damage by worsening ischemic brain damage (7). Cytokines such as IL-1 and TNF are upregulated in stroke and participate in brain inflammation. The secretion of pro-inflammatory cytokines, including IL-1, IL-6, and tumor necrosis factor (TNF) produced by macrophages and microglia, causes brain inflammation (8).
Microglia, which mediate their effect through the CD36 receptor and other molecules, also produce high levels of ROS, which together with pro-inflammatory cytokines, characterize ischemic stroke. These reactions mediate the acute phase of stroke (9). However, it is not yet clear whether microglia influence neurogenesis (10). Non-activated microglia protect the CNS, but on the contrary, after activation, they polarize in M1 cells through TLR4, interferon gamma (IFNg) or granulocyte-monocyte-colony stimulating factor (GM-CSF) receptors, with generation of pro-inflammatory cytokines and arachidonic acid products (11) (Fig. 1).
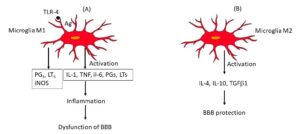
Fig. 1. In panel A we show that the antigen binds TLR4 in Microglia M1 which activates the generation of prostaglandins (PGs) leukotrienes (LTs), and Inducible Nitric Oxide Synthase (iNOS) causing inflammation and dysfunction in the Blood-Brain Barrier (BBB). In panel B the activation of microglia (M2) leads to secretion of anti-inflammatory cytokines which protect the BBB. PGs: Prostaglandins; LTs: Leukotrienes; TLR: Toll-like receptor.
These effects cause a breakdown of the BBB with brain damage. The M2 microglia cells, producing anti-inflammatory cytokines such as IL-10, IL-4, TGFb1 and probably IL-37 and IL-38, tend to dump inflammation with the benefit of the brain system (12) (Table I). The disruption of the BBB can lead to the activation of microglia which are physiologically responsible for the protection and repair of the CNS. The production of IL-10 by the M2 microglia phenotype allows the recruitment of Treg cells, with increased expression of CD86 and MHC-II, counteracting inflammation (13).
Table I. Activation of microglia in response to pathological stressor.

We know that immunological processes contribute to hypertensive phenomena. In particular, activated T cells and macrophages arrive in abundance in the perivascular regions of the arteries and release cytokines such as IL-1, TNF, IL-6, IL-17, and interferon gamma (IFNg), causing vascular resistance and renal fibrosis (14). By causing inflammation, both the innate and adaptive immune systems can provoke damage and malfunction of the organs with the possible consequence of cerebral stroke (15). Microglia, astrocytes, and oligodendrocytes are part of the brain system and play an important role in the formation and healing of ischemic stroke. Thus, they can mediate both harmful and healing effects (16). Oligodendrocyte antigens implicated in neuronal inflammation activate T lymphocytes, involved in the tissue repair process. By producing inflammatory cytokines, active microglia damage the CNS, but in post-ischemic stroke they can generate anti-inflammatory cytokines such as IL-10, IL-37, and IL-38, which regulate the immune response with benefits and recovery of the tissue pathological state (17).
Microglial cells crosstalk with endothelial cells (ECs) contributing to the formation and functioning of the BBB. The CNS is separated from the peripheral circulatory system through the BBB which is composed of cells and biological reactions (18). The BBB, where ECs play a crucial role, protects brain tissue from external insults and keeps the neuronal system safe. Therefore, the BBB is important for the protection of the CNS and the proper functioning of neurons (19). The contact of the ECs of the CNS with other cells such as pericytes and astrocytes, constitutes the BBB and is critical for its efficient functioning. The BBB, with its vasculature, restricts the trafficking of immune cells in the brain, an effect that is impaired when the BBB is disrupted (20). ECs, together with neurons and microglia, form part of the BBB which, when disrupted or dysregulated, leads to the generation of neurological diseases. Activation of ECs in brain tissue induces expression of adhesion molecules ICAM-1 and VCAM-1. ICAM-1 and VCAM-1, and other adhesion molecules such as PECAM-1, participate in the migration of immune cells and, particularly, of CD4+ T cells (22).
Through the interaction with an integrin, the activated CD4+ T cells contribute to the inflammation by crossing the basement membrane of the ECs and in doing so, they can reach the brain tissue (23). The increase in expression of metalloproteins in the inflammatory process contributes to the migration of CD4+ T lymphocytes and neutrophils into the CNS with the secretion of cytokines and chemokines and damage to the glial system. In these biological dynamics, blood vessels play a crucial role in initiating CNS pathologies, including the inflammation that often accompanies these neuropathies such as ischemic stroke (24).
CONCLUSIONS
Therefore, resting microglia cells help to protect the brain tissue, however when they are activated, they can be detrimental to the CNS and may disrupt the BBB, mediating ischemic stroke and hemorrhage (25).
The study of cellular functioning in the BBB allows for better identification of new diagnostic and therapeutic elements which are very important in this field of medicine, where many points are still waiting to be clarified. After a stroke, there can be a loss of BBB that is measurable by various methods such as magnetic resonance imaging, computed tomography, etc (26). Accurate and early examination with these methodologies can prevent intracerebral hemorrhage in individuals presenting with an acute ischemic stroke. Failure of the BBB leads to an accumulation of immune cells in the brain with the secretion of numerous cytokines and growth factors such as VEGF, which are instrumental in causing vascular permeability and neuroinflammation (27).
Thus, after a stroke, the BBB breaks down and fails to function, resulting in the involvement of immune cells that contribute to inflammation and vascular damage. However, more studies are needed to clarify the exact role of pro and anti-inflammatory cytokines in stroke and haemorrhage (28).
Conflict of interest
The author declares that they have no conflict of interest.
REFERENCES
- Schilling M, Besselmann M, Leonhard C, Mueller M, Ringelstein EBernd, Kiefer R. Microglial activation precedes and predominates over macrophage infiltration in transient focal cerebral ischemia: a study in green fluorescent protein transgenic bone marrow chimeric mice. Experimental Neurology. 2003;183(1):25-33. doi:https://doi.org/10.1016/s0014-4886(03)00082-7
- Robida PA, Puzzovio PG, Pahima H, Levi-Schaffer F, Bochner BS. Human eosinophils and mast cells: Birds of a feather flock together. Immunological Reviews. 2018;282(1):151-167. doi:https://doi.org/10.1111/imr.12638
- Khoshnam SE, Winlow W, Farzaneh M, Farbood Y, Moghaddam HF. Pathogenic mechanisms following ischemic stroke. Neurological Sciences. 2017;38(7):1167-1186. doi:https://doi.org/10.1007/s10072-017-2938-1
- Yau B, Hunt N, Mitchell A, Too L. Blood‒Brain Barrier Pathology and CNS Outcomes in Streptococcus pneumoniae Meningitis. International Journal of Molecular Sciences. 2018;19(11):3555. doi:https://doi.org/10.3390/ijms19113555
- Chamorro Á, Dirnagl U, Urra X, Planas AM. Neuroprotection in acute stroke: targeting excitotoxicity, oxidative and nitrosative stress, and inflammation. The Lancet Neurology. 2016;15(8):869-881. doi:https://doi.org/10.1016/s1474-4422(16)00114-9
- Gülke E, Gelderblom M, Magnus T. Danger signals in stroke and their role on microglia activation after ischemia. Therapeutic Advances in Neurological Disorders. 2018;11:175628641877425. doi:https://doi.org/10.1177/1756286418774254
- Jin R, Yang G, Li G. Inflammatory mechanisms in ischemic stroke: role of inflammatory cells. Journal of Leukocyte Biology. 2010;87(5):779-789. doi:https://doi.org/10.1189/jlb.1109766
- Leal MC, Casabona JC, Puntel M, Pitossi FJ. Interleukin-1β and tumor necrosis factor-α: reliable targets for protective therapies in Parkinson’s Disease? Frontiers in Cellular Neuroscience. 2013;7. doi:https://doi.org/10.3389/fncel.2013.00053
- Guruswamy R, ElAli A. Complex Roles of Microglial Cells in Ischemic Stroke Pathobiology: New Insights and Future Directions. International Journal of Molecular Sciences. 2017;18(3):496. doi:https://doi.org/10.3390/ijms18030496
- Valero J, Paris I, Sierra A. Lifestyle Shapes the Dialogue between Environment, Microglia, and Adult Neurogenesis. ACS Chemical Neuroscience. 2016;7(4):442-453. doi:https://doi.org/10.1021/acschemneuro.6b00009
- Medeiros A, Peres-Buzalaf C, Fortino Verdan F, Serezani CH. Prostaglandin and the Suppression of Phagocyte Innate Immune Responses in Different Organs. Mediators of Inflammation. 2012;2012:1-13. doi:https://doi.org/10.1155/2012/327568
- Cherry JD, Olschowka JA, O’Banion M. Neuroinflammation and M2 microglia: the good, the bad, and the inflamed. Journal of Neuroinflammation. 2014;11(1):98. doi:https://doi.org/10.1186/1742-2094-11-98
- Rojo AI, McBean G, Cindric M, et al. Redox Control of Microglial Function: Molecular Mechanisms and Functional Significance. Antioxidants & Redox Signaling. 2014;21(12):1766-1801. doi:https://doi.org/10.1089/ars.2013.5745
- Wang J, Wakeham J, Harkness R, Xing Z. Macrophages are a significant source of type 1 cytokines during mycobacterial infection. Journal of Clinical Investigation. 1999;103(7):1023-1029. doi:https://doi.org/10.1172/jci6224
- Santos Samary C, Pelosi P, Leme Silva P, Rieken Macedo Rocco P. Immunomodulation after ischemic stroke: potential mechanisms and implications for therapy. Critical Care. 2016;20(1). doi:https://doi.org/10.1186/s13054-016-1573-1
- Pekny M, Pekna M. Reactive gliosis in the pathogenesis of CNS diseases. Biochimica et Biophysica Acta (BBA) – Molecular Basis of Disease. 2016;1862(3):483-491. doi:https://doi.org/10.1016/j.bbadis.2015.11.014
- Carniglia L, Ramírez D, Durand D, et al. Neuropeptides and Microglial Activation in Inflammation, Pain, and Neurodegenerative Diseases. Mediators of Inflammation. 2017;2017:1-23. doi:https://doi.org/10.1155/2017/5048616
- Liebner S, Dijkhuizen RM, Reiss Y, Plate KH, Agalliu D, Constantin G. Functional morphology of the blood-brain barrier in health and disease. Acta Neuropathologica. 2018;135(3):311-336. doi:https://doi.org/10.1007/s00401-018-1815-1
- Engelhardt S, Patkar S, Ogunshola OO. Cell-specific blood-brain barrier regulation in health and disease: a focus on hypoxia. British Journal of Pharmacology. 2014;171(5):1210-1230. doi:https://doi.org/10.1111/bph.12489
- Lécuyer MA, Kebir H, Prat A. Glial influences on BBB functions and molecular players in immune cell trafficking. Biochimica et Biophysica Acta (BBA) – Molecular Basis of Disease. 2016;1862(3):472-482. doi:https://doi.org/10.1016/j.bbadis.2015.10.004
- Daneman R, Prat A. The Blood–Brain Barrier. Cold Spring Harbor Perspectives in Biology. 2015;7(1):a020412. doi:https://doi.org/10.1101/cshperspect.a020412
- Lécuyer MA, Saint-Laurent O, Bourbonnière L, et al. Dual role of ALCAM in neuroinflammation and blood–brain barrier homeostasis. Proceedings of the National Academy of Sciences. 2017;114(4). doi:https://doi.org/10.1073/pnas.1614336114
- Sonar SA, Lal G. Differentiation and Transmigration of CD4 T Cells in Neuroinflammation and Autoimmunity. Frontiers in Immunology. 2017;8. doi:https://doi.org/10.3389/fimmu.2017.01695
- Mostofa AGM, Punganuru SR, Madala HR, Al-Obaide M, Srivenugopal KS. The Process and Regulatory Components of Inflammation in Brain Oncogenesis. Biomolecules. 2017;7(2):34. doi:https://doi.org/10.3390/biom7020034
- Kim JY, Kim N, Yenari MA. Mechanisms and Potential Therapeutic Applications of Microglial Activation after Brain Injury. CNS Neuroscience & Therapeutics. 2014;21(4):309-319. doi:https://doi.org/10.1111/cns.12360
- Gilad R, Lampl Y, Eilam A, Boaz M, Loyberboim M. SPECT-DTPA as a tool for evaluating the blood–brain barrier in post-stroke seizures. Journal of Neurology. 2012;259(10):2041-2044. doi:https://doi.org/10.1007/s00415-012-6445-2
- Chiba T, Umegaki K. Pivotal Roles of Monocytes/Macrophages in Stroke. Mediators of Inflammation. 2013;2013:1-10. doi:https://doi.org/10.1155/2013/759103
- Erickson MA, Banks WA. Neuroimmune Axes of the Blood–Brain Barriers and Blood–Brain Interfaces: Bases for Physiological Regulation, Disease States, and Pharmacological Interventions. Dantzer R, ed. Pharmacological Reviews. 2018;70(2):278-314. doi:https://doi.org/10.1124/pr.117.014647